
📼 游戏介绍
游戏特色
欧美SLG/赛博朋克 赛博穿越指南 第二季 Cyberstray 202 / /690M
2026-07-19 电脑游戏
游戏介绍 更新日志
你将继续扮演那位被命运选中的流浪者,探索都市中的阴谋与霓虹灯光下低语的陌生人。 新加入的角色带来更复杂的盟友与敌对关系,而神秘的穿越现象背后隐藏着更深的真相。 你将如何在浪漫邂逅与致命危机中崛起?是掌控命运,还是被情感吞噬?一切由你决定。
游戏截图
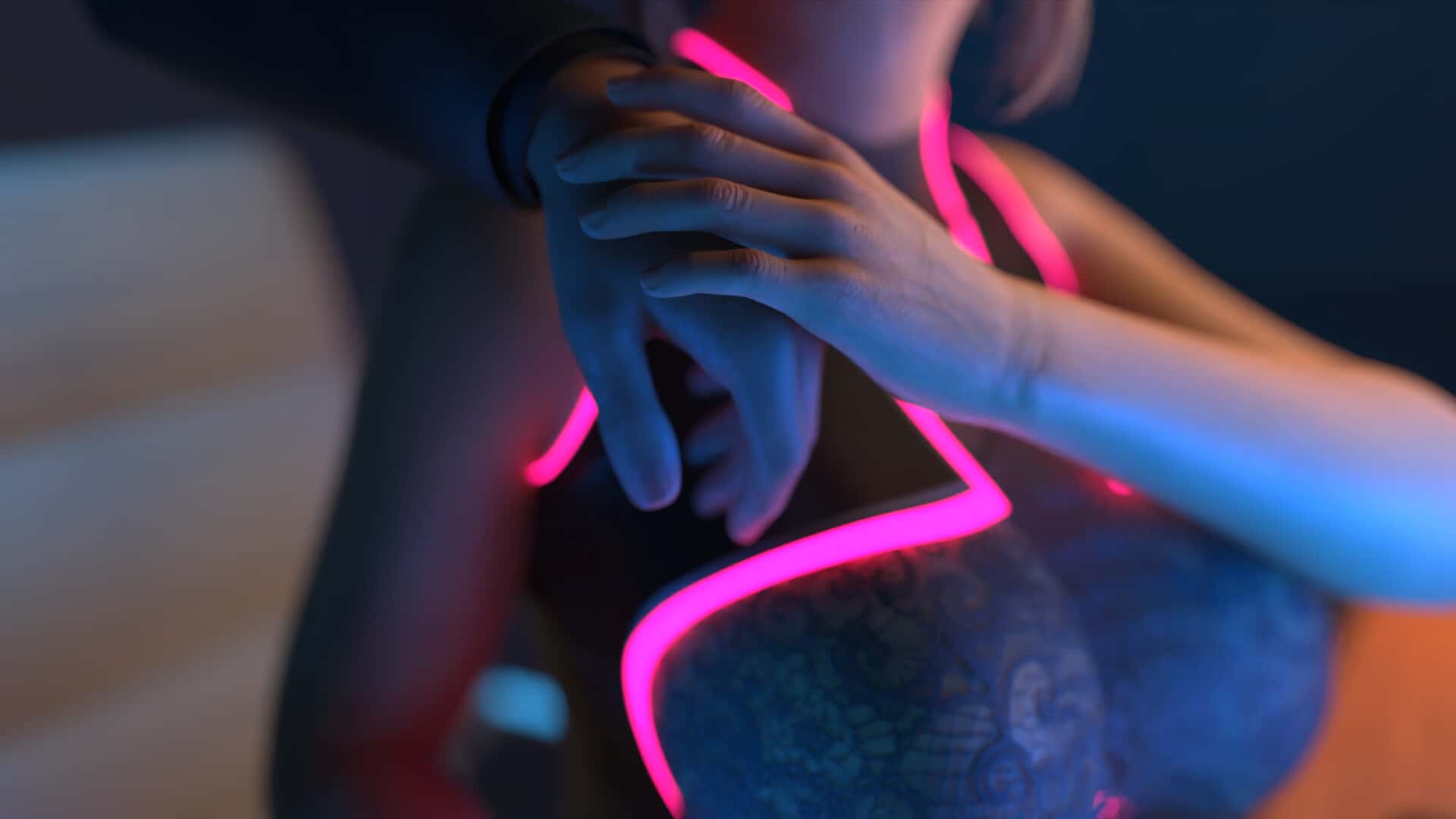


🎵 游戏攻略
N/A